二重構造説
日本人は、日本人の起源論が好きである。実際、日本人の系統は単純ではなく、いまだに詳細がわかっていない。
民族集団の系統関係と言語の系統関係がよく一致することから、言語の類似性から人種や民族の系統を探る試みが続けられてきた。言語学的にみると、韓国・朝鮮語と日本語は、いずもアルタイ諸語との関係が指摘されているが、確かな類縁関係は示されず、言語として孤立している。さらにオホーツク海の周囲には、アイヌ、ニブフ(ギリヤーク)という孤独語が分布している。これだけ孤立した言語が並んでいる地域は珍しい。
日本人の系統は単一ではなく、先住の縄文人が狩猟採集生活を行っていたところに、稲作技術を持った弥生人が渡来し、混合したのが日本人だというのが二重構造説である。この二重構造説によって、日本人が単一の系統では説明できないことが解釈できる。弥生文化が伝わらなかった北海道では縄文文化が残り(続縄文文化)、オホーツク文化と交流し、現在のアイヌに引き継がれた、あるいは、弥生文化が遅れて伝わった沖縄にも、縄文人の要素がより多く残っているという説とも整合性がある。
二重構造説では、縄文語は南方系の、おそらくはオーストロネシア系であり、それにアルタイ系の弥生語が流入して混じり合ったものが日本語だとされるが、文字による記録のない過去の言語を推測するのは難しい。
おそらく、日本列島は大陸の辺縁にあって、移住してきた集団の「吹きだまり」になりやすく、しかし海によって地理的に隔離されているので、人々が外部から渡来するよりも、内部で混合するほうが速く、異質なものが混ざって表面上は均質になりやすかったのではないかと考えられる。
よく知られたSNP
アルコールの代謝
遺伝子をDNAレベルで分析するのが難しかったころには、表現型がはっきり現れ、しかも遺伝の様式が単純であるSNP(一塩基多型)を用いた分析が行われてきた。ALDH2(アセトアルデヒド脱水素酵素)は、エチルアルコールの分解に主要な役割を果たす酵素の遺伝子で、このALDH2上に存在するSNPであるrs671の、G/Gはアルコール分解能力が高く、A/Gは中間で、A/Aはアルコール分解能力が低い。Aは東アジアに特有の変異である。


東アジアにおけるA/AとG/Aの頻度[*2]。
日本人全体におけるG/Gは56%で、逆にA/Aは4%である。(これを書いている蛭川はA/Aである。)Aは本土の西日本に多く、また揚子江(長江)下流域に多い。揚子江下流域は水稲耕作とアルコール飲料の発祥の地だと考えられているが、ここから弥生文化が西日本に伝わったと考えられる。

なお、アルコールを分解する酵素であるADH1Bの分布も麦と水稲耕作の広がりと対応している。
ADH1B * 47His対立遺伝子頻度の分布[*4]

アルコールを分解しやすいADHの突然変異と、アセトアルデヒドを分解しにくいALDHの突然変異の両方があると、アセトアルデヒドが蓄積しやすく「酒に弱い」体質になるが、これが酒の伝播と関係しているのは、「酒に強い」、問題行動を起こしたりアルコール依存症になる遺伝的気質に対して淘汰圧がかかる遺伝子・文化の共進化が起こったとも解釈できる。
耳垢
もうひとつ、よく知られたSNPが、耳垢(湿型、乾型)である。ABCC11遺伝子上のSNPであるrs17822931がG/GまたはG/Aだと湿型になり、A/Aが乾型となる。
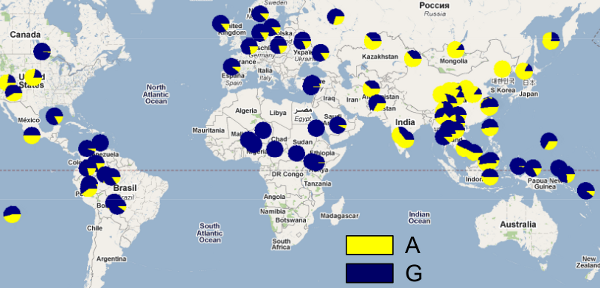
AとGを独立に集計した耳垢の変異の分布[*5]。
乾型の遺伝子であるAは、アフリカで全くみられず、中央アジア、北東アジア、北米に多い。
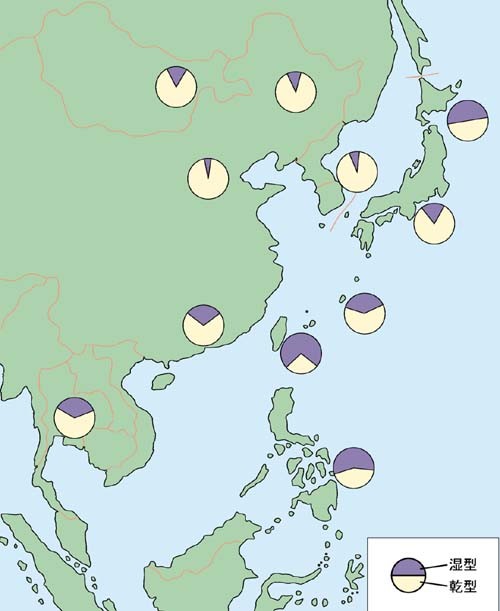
東アジアにおける耳垢タイプの分布[*6]。
日本列島付近では、大陸の北岸にはA(乾型)が多く、東南アジアにはG(湿型)が多い。(これを書いている蛭川はA/Aで、乾型耳垢である。)このことから、湿型のGを多く持った南方系の縄文人の集団に、乾型のAを持ったアルタイ系の弥生人が、華北または朝鮮半島から渡来し、アイヌや沖縄に縄文人の遺伝子が多く残ったというストーリーが描ける。
Y染色体ハプログループ
分子遺伝学の進歩により、表現型やSNPではなく、ある程度まとまったDNAの塩基配列が分析できるようになってきた。似たような遺伝子群を持つグループは、ハプログループとして分類される。

約7万年前にアフリカの祖先と分岐したとされる、最も古いモンゴロイドのDは、主にアンダマン諸島、チベット、日本(アイヌ・縄文・沖縄系)に分布する。Dは最初に東アジアに拡散したグループで、後から来た集団が流入した後、島嶼や高山地帯にわずかに残ったと考えられる。
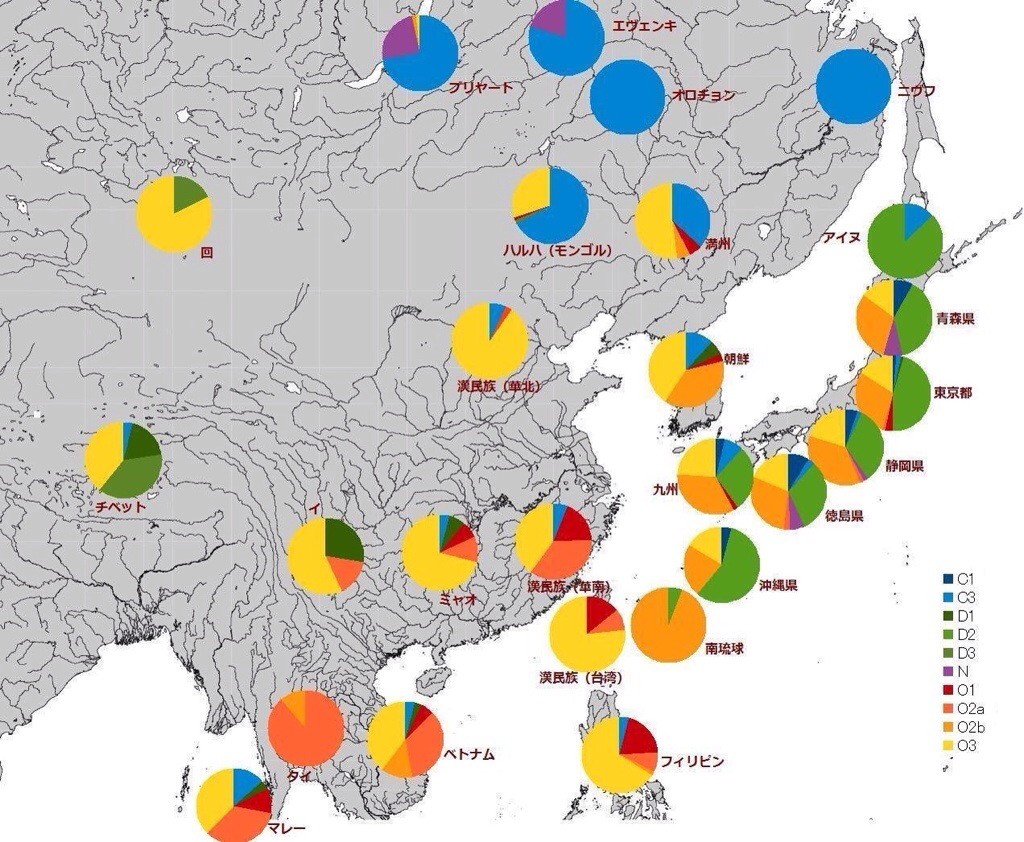
東アジアにおけるY染色体ハプログループの分布[*8]。現在、D1とD2は、D1a、D1bと呼称が変わっている。
東アジアでは、非常に古いモンゴロイドであるDの遺伝子が分布しているのはチベットと日本だけである。これが日本の先住民である縄文人で、そこにO型の遺伝子を持つ弥生人が渡来したとすると、二重構造説が支持される。
父系でのみ伝達されるY染色体の遺伝子は、男性が主となる大規模な移住の結果を反映しやすい。男性のほうが長距離を移動しやすく、少数の男性が多数の女性との間に子を残しやすいからである。このような視点からみて、本州でも半数の集団がDを保持しているのは、渡来した弥生人の数が少ないこと、弥生人が縄文人を征服したわけではないことを示している。じっさい、縄文時代から弥生時代にかけて、大規模な戦いが起こったという考古学的証拠はない。
Y染色体ハプログループの分布をより細かくみると、アイヌと沖縄でDの割合が高く、またCが北方に多いことから、アイヌ人の成り立ちにはオホーツク海の人々の影響があることもわかる。CはDについで古い古モンゴロイドで、北東シベリアに残った人々は古アジア諸語という言語の話者である。先島(南琉球)は考古学的には縄文文化の外部で、オーストロネシア系だという仮説も、ある程度は支持される。
なお、この文章を書いている蛭川のY染色体のハプログループはD1b1aで、縄文系である。低いアルコール分解能力と乾型耳垢だということもあわせて考えると、東南アジア由来の縄文人と渡来系弥生人の混血らしい。これは、日本人としては一般的なパターンである。

蛭川の染色体。大部分が日本人の系統(暗い茶色)だが、9、13、18、22番染色体の片方が韓国・朝鮮系(明るい茶色)で、19番染色体の片方が漢民族の系統[*9](橙色)だと推測されている。
ミトコンドリアDNAハプログループ
ミトコンドリアDNAは核のDNAと異なり、ゲノムのサイズが小さく、古人骨の解析も容易だという特徴がある。父系で遺伝するY染色体とは対照的に、母系遺伝なので、男性の移住よりも規模の小さい移住も反映される。

ミトコンドリアDNAハプログループの分布と推定される拡散経路[*10]。
人種[*11]の分類

東アジアにおけるミトコンドリアDNAパプログループの分布[*12]。
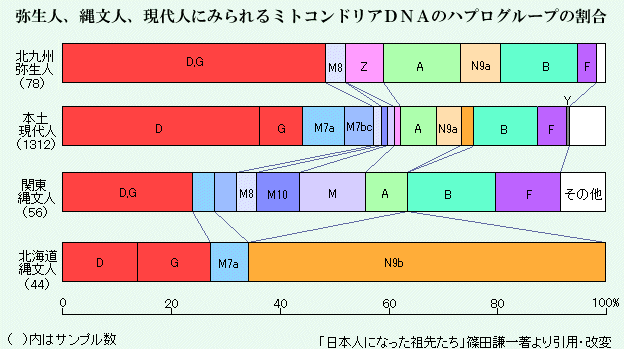
日本列島における現代人と古人骨のミトコンドリアDNAハプログループの割合[*13]。
ミトコンドリアDNAのハプログループはY染色体よりも小規模の移住を含むので、解像度は高いが、そのぶん複雑に入り組んでおり、解釈が難しくもある。本州の日本人・沖縄人でいちばん多いのがD・Gで、これは渡来系弥生人の半分を占めるグループであり、関東の縄文人、北海道の縄文人、北海道アイヌでもある程度みられる。これらから、おもにD・Gは、北方に由来する渡来系弥生人に由来すると推測される。
しかし、本州の日本人、沖縄人、そして関東の縄文人の集団は多様であり、D・G以外にも多数のハプログループが存在し、土器の様式などでは一様性があった縄文人がもともと多民族的な集団だったことをうかがわせる。Bはオーストロネシア語族と関連し、Mはオーストロアジア語族と関連していると推測されている。またFは東南アジアに広くみられるが、特定の言語との関係は不詳である。またAは古アジア諸語と関連している。古アジア諸語の話者が、がかなり古い時代に東南アジアから北上し、ベーリング地峡を越えて北米にまで到達したということを考えると、縄文時代の日本列島民はすでに、かなり古い時代から新しい時代にかけて、東南アジア方面から渡来した人たちからなる、重層的な集団であったらしい。(日本列島への人類の移住は3万8000年前で、縄文時代の始まりは1万5000年前、弥生時代の開始が2300年前だと推測されている。)
北海道縄文人と北海道アイヌには、本州以南と違ってB、F、Aがみられず、その代わりにY(N9)が多い。Yはオホーツク海周辺の民族で高頻度にみられる型で、アイヌ文化がオホーツク文化と関連することを支持しているが、北海道の縄文人は縄文時代から本州の集団とは異なっていたとすると、この点でも縄文人は多様であったといえる。
なお、これを書いている蛭川のミトコンドリアDNAのハプログループはBやFの共通祖先のR(おそらくR11)である[*14]。アフリカを出たL3の集団が西南アジアに到達し、約6万年前に南北に分かれて南インドに向かったのがMで、北インドに向かったのがN→Rである。RからはHV(ヨーロッパ)、JT(アフロアジア)、F(東南アジア、日本にも多い)、B(オーストロネシア、日本にも多い)、P(東ヘスペロネシア・ニューギニア・オーストラリア)、U(アフロアジア)が分岐した。これらの集団が分岐する以前の古いRの遺伝子はヨーロッパからオーストラリアに至る地域に断片的に残存しているが、とくに多いのはNからRが分岐した北インドである。
(個人で申し込める遺伝子検査についてはこちらを参照のこと。)
記述の自己評価 ★★★☆☆
(講義メモとして作成したページで、議論の精度はやや荒く、引用も内容・形式ともに不十分ですが、当面、このページは残します。一部分は加筆修正して蛭川研ブログ新館に移転する予定です。)
2016/12/05 JST 作成
2019/10/07 JST 最終更新
蛭川立
*1:「Alcohol flush reaction」『Wikipedia』(2022/02/16 JST 最終閲覧)
{kind=link}
*3:Masayuki Sakiyama, Hirotaka Matsuo, Airi Akashi, Seiko Shimizu, Toshihide Higashino, Makoto Kawaguchi, Akiyoshi Nakayama, Mariko Naito, Sayo Kawai, Hiroshi Nakashima, Yutaka Sakurai, Kimiyoshi Ichida, Toru Shimizu, Hiroshi Ooyama, and Nariyoshi Shinomiya (2017). Independent effects of ADH1B and ALDH2 common dysfunctional variants on gout risk. Scientific Reports, 7(1), doi: 10.1038/s41598-017-02528-z.
*4:Hui Li, Namita Mukherjee, Usha Soundararajan, Zsanett Tárnok, Csaba Barta, Shagufta Khaliq, Aisha Mohyuddin, Sylvester L. B. Kajuna, S. Qasim Mehdi, Judith R. Kidd, Kenneth K. Kidd (2007). Geographically Separate Increases in the Frequency of the Derived ADH1B*47His Allele in Eastern and Western Asia. American Journal of Human Genetics, 81(4), 842-846.
*5:Razib Khan (2010). 「Body Odor, Asians, and Earwax」『Discover』(2022/02/16 JST 最終閲覧)
*6:九州大学総合研究博物館「縄文人と弥生人」『九州大学ミニミュージアム 倭人の形成 九州大学古人骨資料からみた日本人の形成』(2022/02/16 JST 最終閲覧)
*7:Human mitochondrial DNA haplogroup (Wikipedia)
*8: 青海波 (2014).「DNAで見る日本人 遺伝子の情報」『古代史俯瞰 by tokyoblog』(2022/02/16 JST 最終閲覧)
*9:「漢民族」もまた、中華帝国とその周辺の諸民族の混血だと考えられている。
*10:「Human mitochondrial DNA haplogroup」『Wikipedia』(2022/02/16 JST 最終閲覧)
*11:「人種 race と」いう概念は、しばしば差別的なニュアンスを含む。人間集団に遺伝的な変異があるのは事実だが、ここでは差別的なニュアンスは含めていない。
*12:篠田謙 (2007).『日本人になった祖先たちーDNAから解明するその多元的構造ー』NHK出版.
*13:篠田謙 (2007).『日本人になった祖先たちーDNAから解明するその多元的構造ー』NHK出版.
*14:Rは本土の日本人で0.1%しか存在しない。Tanaka, M, Cabera, V. M. et al. (2004).